
Bone
- teachanatomy

- Apr 12
- 18 min read
Updated: Apr 14
Bone is a specialized form of connective tissues. It is made up of cells, ground substance and fibers. It is also known as the osseous tissue. The ground substance and the fibers form together the bone matrix. The fibers are collagen fibers. The ground substance is mineralized. The collagen fibers are type 1 collagen fibers that are regularly arranged. The regular arrangement of collagen fibers imparts a lamellar appearance to the matrix. Fibers in each lamella lie parallel to each but are perpendicular to those in the neighboring lamellae

Bone Cells
There are four types of bone cells:
Osteoprogenitor: These are also known as osteogenic cells and bone lining cells. They are relatively undifferentiated cells that originate from mesenchymal cells and are capable of proliferation and development into osteoblasts. A vascular environment is a necessary condition for osteoprogenitor cells to differentiate into osteoblasts. A high oxygen tension (↑O2) is needed for this process. Osteoprogenitor cells are present in both the periosteum where they are called periosteal cells, and in the endosteum where they are known as endosteal cells. Osteoprogenitor cells are spindle-shaped cells that may show tiny thin processes known as filopodia. They play important functions in bone development (osteogenesis) and bone repair (bone healing). They are thus more frequently seen in developing and healing bones. Their numbers are capabilities for producing osteoblast are reduced at old age.
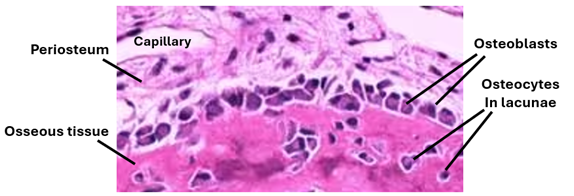
Osteoblasts: These are fully differentiated cells that develop from osteoprogenitor cells under the influence growth factors. During bone development and growth, they synthesize and secrete bone matrix proteins such osteocalcin. They also synthesize and secrete collagen type 1 which constitutes more than 90% of bone matrix proteins. Osteoblasts are large cells with a basophilic cytoplasm, eccentric nucleus and a cell few processes. During bone development, they arrange themselves in sheets that cover the bone surface. In histological sections, they may resemble simple cuboidal epithelium. Their differentiation requires a good blood supply. In the absence of an adequate blood supply, chondroblasts and cartilage are formed. Osteoblasts have another important function which is mineralization of the matrix they secrete. They synthesize and secrete alkaline phosphatase which mineralizes the matrix. The unmineralized bone matrix is called osteoid. It is a basophilic amorphous material made of the ground substance and collagen types 1 fibers. Meanwhile, the processes of osteoblasts elongate and become extensive. Ultimately, the osteoblasts and their processes are surrounded by the newly formed bone matrix. Then, the osteoblasts become osteocytes.
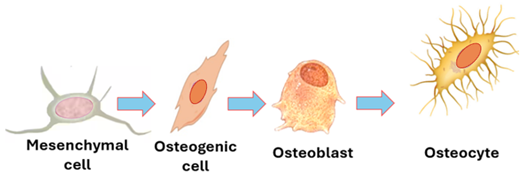
Osteocytes: Once entrapped within osseous tissue they secrete, osteoblasts are known as osteocytes. Their bodies occupy potential spaces known as lacunae and their processes occupy tiny channels known as bone canaliculi. Osteocyte processes are slender and numerous; they adhere to each other, and communicate via gap junctions. Osteocytes function in maintaining the integrity of bone; they continue to secrete components of the bone matrix for maintenance and repair. They also send signals to osteoblasts and osteoclasts modulating their functions. In addition to osteocyte processes, the canaliculi contain tissue fluids. The lacunae containing the bodies of osteocytes also contain tissue fluids. Tissue fluids from periosteal vessels irrigate osteocytes of the entire bone via interconnecting bone canaliculi and lacunae. Osteocytes are long living cell (10-20 years) that are terminally differentiated. They are stellate cells that appear smaller than osteoblasts. The cytoplasm of osteocytes contains few organelles.
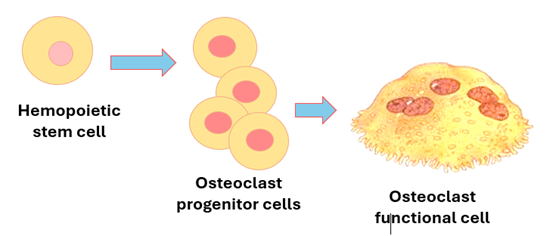
4. Osteoclasts: The developmental origin of osteoclasts is different from that of osteocytes. Osteoclasts develop from hemopoietic stem cells when appropriately stimulated. They differentiate into monocyte / macrophage progenitor cells, then into osteoclast precursor cells, then into functional osteoclasts. Several osteoclast precursors join each other to form a single osteoclast. Osteoclasts are huge cells measuring about 200um in diameter. They are multinucleated cells often containing four or more nuclei. The cytoplasm contains lysosomes, vacuoles and vesicles that reflect the high activity of these cells in phagocytosis. The cytoplasm also contains rER and well-developed Golgi complexes both needed or formation of lysosomes. Osteoclasts break down bone tissue by a process called bone resorption, which is quite important for maintenance, repair and remodeling of bone. Osteoclasts secrete collagenase, gelatinases and other proteases which break down collagen and organic matter. In addition, they secrete acid phosphatase and hydrogen ions that dissolve matrix minerals. By doing so they help in regulating the blood calcium levels. Osteoclasts are stimulated by the parathyroid hormone and suppressed by calcitonin which is produced by the thyroid gland. Osteoclasts reside on bone surfaces within bays called Howship’s lacunae. When active they erode the bone surface and dig in forming Howship’s lacunae. The process of bone resorption starts by acid secretion which dissolves minerals. Then there is secretion of the proteolytic enzymes which digest the demineralized matrix. They then phagocytose the degraded matrix fragments. The characteristic raffled border of osteoclasts plays an important role in the process of phagocytosis. Osteoclasts belong to a body system made of phagocytic cells called the monocyte / phagocyte system.
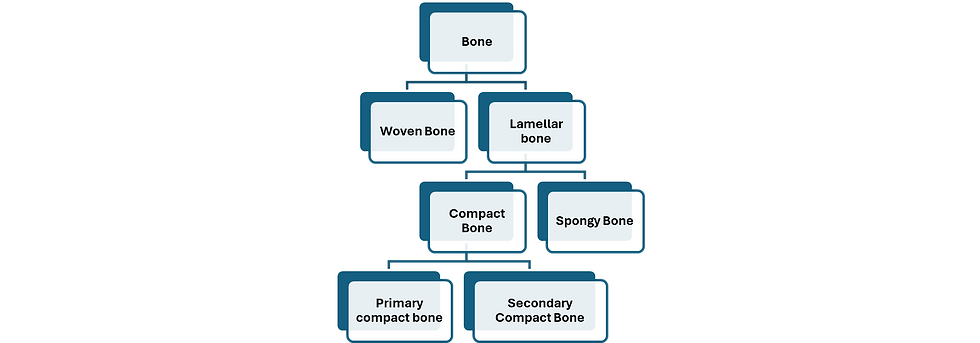
Classification of Bone
Bone tissue is classified into woven bone and lamellar bone. Woven bone is immature and lacks lamellae. Lamellar bone is a mature bone. It is characterized by clear lamellae. It is of two types: lamellar compact bone and lamellar spongy bone. Compact bone is either primary or secondary
Woven Bone
Woven bone is an immature bone present in the skeleton of young growing children aged less than 5 years, or around bone fracture sites. It appears disorganized because it is laid down rapidly. It does not form on existing bone or cartilage. Woven bone is characterized by an abundance of osteoblasts and osteocytes. It is less dense than mature bone. Its collagen fibers are thick and loosely packed and are disorganized and accordingly it does not show distinct lamellae. Woven bone is weaker and less rigid than mature bone and can become highly mineralized and brittle. Its osteocytes and lacunae are usually larger than those of mature bone.
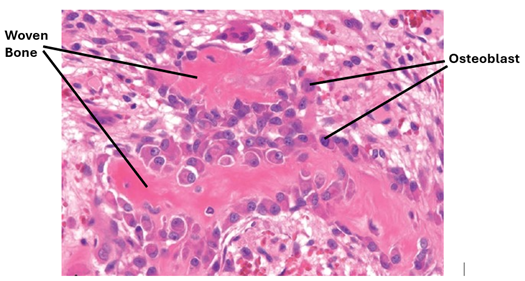
Lamellar Bone
Lamellar bone is mature bone. It has well organized layers called lamellae. Lamellar bone is of two types: spongy bone and compact bone. Compact bone is of two types:
1. Primary compact bone is the compact bone which is formed first. It is a well-organized bone made only of Haversian systems.
2. Secondary compact bones replace the primary compact bone during remodeling. It is also well organized but contains interstitial lamellae in addition to the Haversian lamellae.
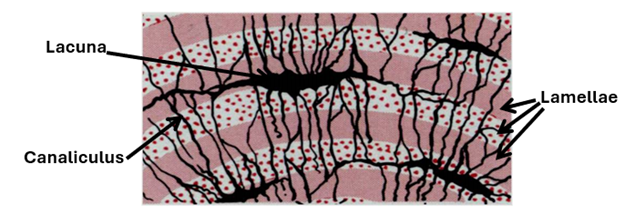
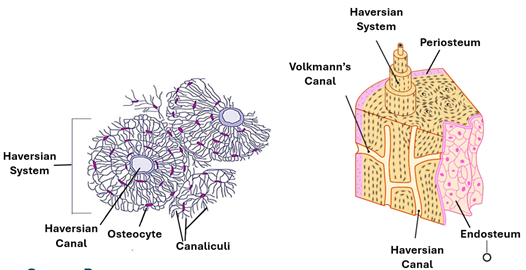
Lamellae, Lacunae and Canaliculi
Mature bone tissue is characterized by lamellae, canaliculi and lacunae. Lamellae are layers of osseous tissue arranged parallel to each other. The lamellae are wavy in spongy bone and concentric circular in compact bone. The regular arrangement of collagen type 1 fibers is the cause of bone lamellation; parallel to each other in each lamella and perpendicular to fibers in the adjacent lamellae. Lacunae are the spaces occupied by osteocytes. They are potential spaces around which the osteoblasts and osteocytes have laid down the bone matrix. The lacunae are interconnected by tiny channels called canaliculi, which facilitate passages of tissue fluids containing nutrients the periosteal blood vessels to all osteocytes within the osseous tissue. The canaliculi contain the slender long processes of osteocytes.
Compact and Spongy Bone
Both compact and spongy bone are lamellar bone but have different porosities. Compact bone is also known as cortical bone. It is quite dense with a porosity of 5-10%. It is found primarily in the shaft (diaphysis) of long bones and forms the outer shell around spongy bone in other locations. Haversian systems are the characteristic structural feature of mature compact bone. The Haversian system, which are also known as osteons, are cylindrical bony structures. Spongy bone, which is also called cancellous bone and trabecular bone is much more porous with porosity of 50-90%. It is found in the epiphysis of long bones, in the vertebrae and in the pelvis and other flat bones of the body. Trabeculae are the characteristic structural feature of spongy bone; they are platelike bony structures.
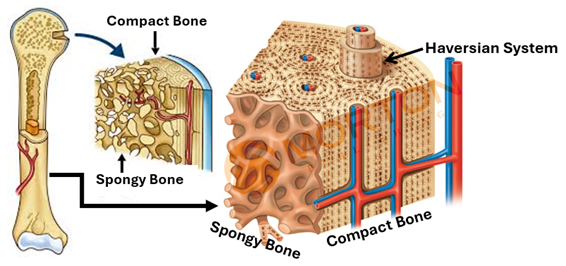
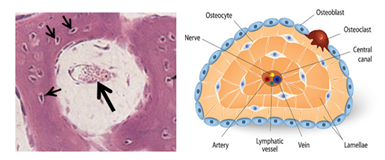
Compact Bone
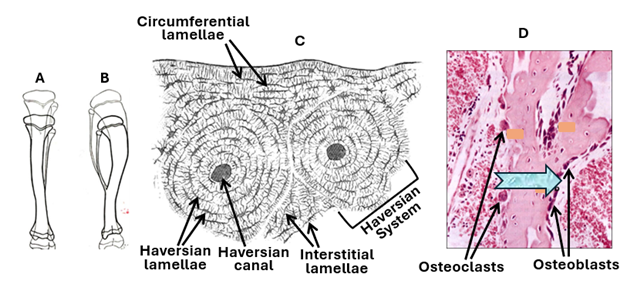
Compact bone is characterized by Haversian systems, which are also known as osteons. Compact bones contain numerous closely packed Haversian systems. Each system (osteon) is a cylindrical structure consisting of a canal called the Haversian canal surrounded by many concentric lamellae known as the Haversian lamellae. The Haversian canal contains blood vessels that supply osteocytes and other bone cells with oxygen and nutrition. The canal also contains nerve fibers and osteogenic cells known as endosteal cells. Haversian systems of compact bone are traversed by horizontal or oblique canals called Volkmann's canals. Volkmann’s canals carry blood vessels and nerves from the periosteum into the substance of the compact bone. Circumferential lamellae long lamella that run parallel to each other and the bone surface underlying the periosteum, They imapart a smooth texture to the outer surface of bones.
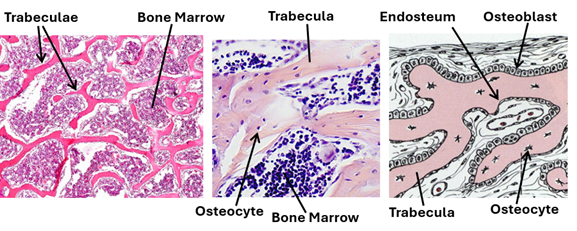
Spongy Bone
Spongy bone, which is also known as cancellous bone, resembles compact bone in that it too has osseous lamellae, lacunae, osteocytes and canaliculae. However, the lamellae of spongy bone are arranged parallel to the surface, not concentrically arranged around a small canal. The canaliculi connect lacunae to each other and to the periosteum, there are no Haversian and Volkmann’s canals. Spongy bone is characterized by bone tissue plates known as trabeculae, which branch in an irregular manner. The trabeculae pass in different directions forming a three-dimensional latticework. Although irregularly arranged, the spongy bone trabeculae provide maximum strength to the bone. The trabeculae surround irregular spaces known as marrow cavities because they contain bone marrow. Marrow cavities are not present in compact bone, and that is why the spongy bone is lighter than compact bone. It has a porosity higher than that of compact bone. The marrow cavities may constitute up to 75% of spongy bone volume. It is to be noted that both the osseous tissue of the spongy bone and the bone marrow it contains originate from mesenchyme. The surfaces of spongy bone trabeculae are covered by endosteum. During bone development osteoblast are present on the surfaces of these plates. Cancellous bone is present in flat and irregular bones and the ends of long bones.
Structure of Long Bones
Long bones have a shaft known as the diaphysis and two enlarged ends, each known as the epiphysis. The diaphysis and the epiphysis are joined together by the metaphysis. The diaphysis is mainly compact bone. The epiphysis and the metaphysis is composed of spongy (cancellous) bone covered by a shell of compact bone. The epiphyseal compact bone covered by a thin layer of articular hyaline cartilage. The junction between the epiphysis and the metaphysis is called the epiphyseal plate. In growing bones, the epiphyseal plate is a proliferative plate of hyaline cartilage. This is the region of bone that grows longer before and during puberty.
Bone Development (Osteogenesis)
Bone originates from the embryonic mesenchyme either directly or via cartilage. Direct bone development is the method whereby flat bones of the skull are formed; it is called intramembranous ossification. Axial and appendicular bones are formed by the indirect method of osteogenesis which is called endochondral ossification.
Intramembranous Ossification
In this type of osteogenesis mesenchymal cells condense forming a highly vascularized sheet. Within this highly vascular environment the mesenchymal cells transform into osteogenic cells then into osteoblasts. Osteoblasts secrete osteoid. Osteoid is an unmineralized bone matrix made of the ground substance and type 1 collagen fibers. Osteoblasts then secrete alkaline phosphatase which mineralizes the osteoid transforming it into osseous tissue. The osteoblasts develop cytoplasmic processes and become entrapped in the matrix. When fully entrapped in the matrix they become known as osteocytes. Collagen fibrils are at first irregularly arranged forming woven bone.
Both spongy and compact bone can develop by intra-membranous ossification. When osteoblasts lay bone matrix in concentric layers around blood vessels, compact bone develops. If they lay down matrix into irregular trabeculae, spongy bone is formed. Irregular trabeculae do not mean woven bone; woven bone is characterized by irregularly arranged collagen fibers.
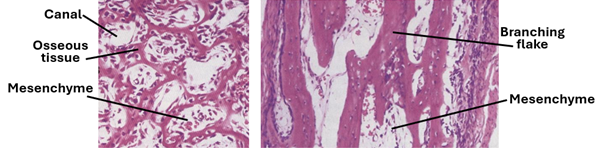
Development of Haversian Systems
Haversian systems develop around blood vessels within the mesenchyme. Initially, each developing Haversian system shows a large canal full of mesenchyme containing a blood vessel. The canal gradually becomes narrower as more osseous tissue is deposited in concentric layers around the canal by the osteoblasts present on the surface. Young Haversian systems have smooth wide Haversian canals; old ones have narrow canals.
Endochondral Ossification
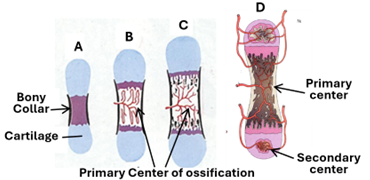
In this type of bone development, the mesenchyme condenses and develops into cartilaginous models that foreshadow the bone shape. The model is made of hyaline cartilage. The length and diameter of the model increase due to both appositional and interstitial growth of the cartilage. First blood vessels invade the perichondrium (the future periosteum) of the model’s shaft stimulating development of osteoprogenitor cells and osteoblasts. As a result, a bony collar called the subperiosteal bone collar is formed. During formation of the bone collar, chondrocytes hypertrophy, and the cartilage matrix becomes narrower and calcifies. Then, periosteal buds made of blood vessels and accompanying mesenchymal cells pass into the calcified cartilage forming the primary center of ossification, which is also called the diaphyseal center of ossification.
The Primary Center of Ossification
Within the vascular environment of the primary ossification center mesenchymal cells develop into osteoblasts that lay osseous tissue on the surface of calcified cartilaginous matrix. The process of ossification proceeds in both directions along the diaphysis towards the metaphysis.
The Secondary Centers of Ossification
These centers develop after birth. Blood vessels and mesenchymal cell invade epiphyseal cartilage. At the same time chondrocytes hypertrophy and the matrix calcifies. In this vascular environment the mesenchymal cells develop into osteoblasts and hemocytoblast. The osteoblasts lay osseous tissue whereas the hemocytoblasts differentiate into marrow formation. The secondary centers enlarge leaving the hyaline cartilage only at the articular surface of the epiphysis and epiphyseal (growth) plate in the diaphysis.
Bone Growth and Remodeling
Bones are rigid structures. Nonetheless, they can grow and change shape. The increase in the size of flat bones and the width of long bones is brought about by appositional growth facilitated by the periosteal osteoprogenitor cells. New bone tissue (subperiosteal bone) is laid beneath the periosteum by the osteoblasts that differentiate from the osteoprogenitor cells present in the deep regions of the periosteum. This process is reminiscent of intramembranous ossification. On the other hand, the increase in the length of long bones is accomplished by interstitial growth of the epiphyseal plate cartilage and the subsequent deposition of bone on the calcified cartilage. This process is reminiscent of endochondral ossification.
The Epiphyseal Plate
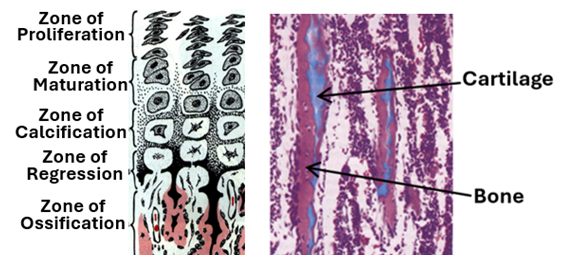
The epiphyseal plate is a distinct plate present between the diaphysis of long bones and each of the two epiphyses of long bones in young people. It is present only during the growth period of an individual and vanishes (epiphyseal closure) soon after puberty. In adults, the plate is replaced by an epiphyseal line. Epiphyseal closure usually occurs at the age of 14-15 years in girls and the age of 15-17 years in boys. The epiphyseal plate is characterized by distinct histological zones that correspond to the different stages of development and regression of the plate cartilage. These zones are:
1. The first zone of the plate which is present at its epiphyseal end is the zone of reserve.
2. Beneath the zone of reserve is the zone of where chondrocytes proliferate by mitosis under the influence of growth hormone and become closely arranged in columns.
3. The zone of maturation is present beneath the zone of proliferation. In this zone cell divisions cease, and the chondrocytes enlarge.
4. The zone of hypertrophy comes next and is often inconspicuous. In this zone chondrocytes hypertrophy and become larger.
5. In the zone of calcification, calcium is deposited on the cartilaginous matrix.
6. Calcification of the cartilage matrix prevents diffusion of tissue fluids and nutrients to chondrocytes causing degeneration chondrocytes and the regression of cartilage. Accordingly, this zone is called the zone of regression.
7. The zone of ossification is the zone closest to the diaphysis. In this zone new primary cancellous bone is formed on top of regressing cartilage.
These zones are essentially reflections of chondrocyte proliferation, differentiation, death and new bone formation. Under the influence of the growth hormone and other growth factors, chondrocytes differentiate from quiescent round cells into proliferating flattened cells to large fully differentiated cells. Calcium is then deposited causing chondrocyte death.
Remodeling
Throughout the individual life, bones change size and shape. This is brought about by modeling and remodeling. Before birth bones are formed and modeled. The change in shape or structure of modeled bone is called remodeling. Remodeling occurs throughout life and is a process of rearrangement of internal bone organization to accommodate for change in bone shape and pressure forces. It also facilitates replacement of old bone tissue with a new one. Remodeling is achieved by the combined activities of osteoclasts and osteoblasts. Osteoclasts remove old bone tissue in regions and osteoblast add new bone tissue in other regions. Remodeling replaces small amounts of primary or old bone with new secondary bone at a time. During remodeling of compact bone, Haversian canals first become wider, and irregular due to removal of osseous tissue by osteoclasts, then the canals once again become narrower and regular due to deposition of new osseous tissue osteoblasts. Interstitial lamellae and reversal (cement) lines are remnants of old Haversian systems present between the newly formed Haversian systems; they indicate remodeling. Remodeled bone tissue characterized by interstitial lamellae is known as secondary bone; the primary bone tissue lacks interstitial lamella and cement lines. Cement lines hold old and new bone tissues firmly. Cement lines (reversal lines) are thin layers of glycoprotein. Remodeling serves several functions: It repairs damaged bone, prevents accumulation of too much old bone, and is a means for storing excess calcium and phosphorus in the blood and supplying the body with these two minerals when they are needed. Calcium removal and deposition is controlled by calcitonin and parathormone. Remodeling renews the entire adult skeleton every ten years.
Joints
Joints are regions where ends of two or more bones meet; they are structures that join adjacent bones together. They are also known as articulations. Most joints are mobile, allowing a degree of movement of the articulated bones. Joints are of different types. They could be fibrous, cartilaginous or synovial. Fibrous joints could be sutures as in the skull, or syndesmosis as in the interosseous membrane of the forearm. Cartilaginous joints could be primary hyaline cartilage joints as in the sternocostal joints, or secondary fibrocartilage joints as in the intervertebral joints and the pubic symphysis. Synovial joints have several functional and anatomical subdivisions e.g. ball and socket, plane, pivot, hinge, saddle and condylar.
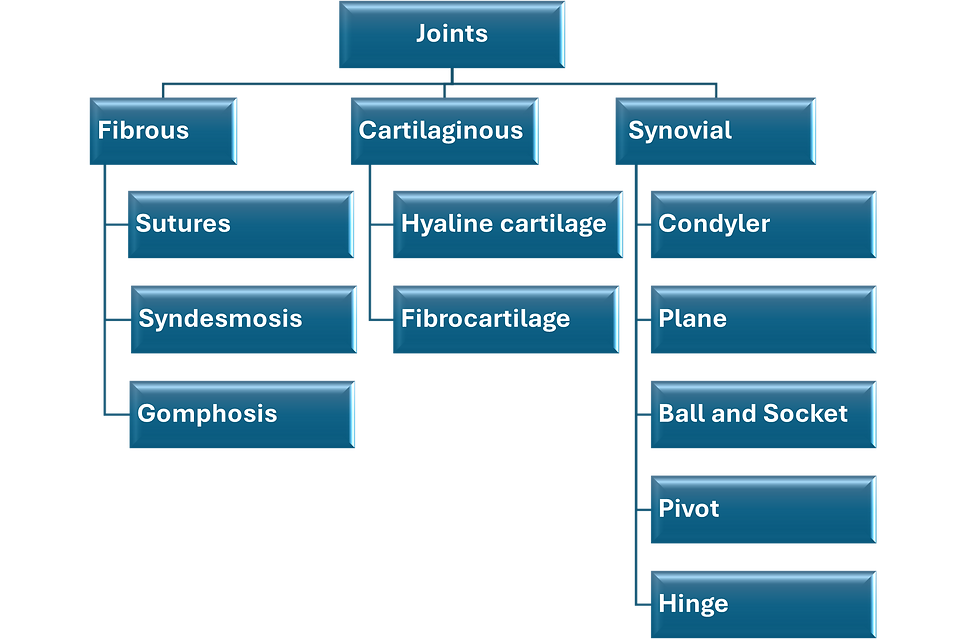
Fibrous Joints
Fibrous Joints, which are also known as synarthroses are where connective tissues unite the ends of two bones together. They lack a synovial cavity and are usually immovable. They are of three subtypes called sutures, syndesmoses and gomphoses. Sutures are found only in the cranium joining the plate-like cranial bones together. The loose connective joining the bones together in infants allows bone growth and slight degree of mobility. The spaces between the cranial bones occupied by the joining loose connective tissue are called fontanelles. As the child grows and the cranial bones become larger the loose connective differentiates into a tough fibrous connective tissue and eventually replaces by bone. Ultimately the cranial bone merge into a single bone. Syndesmoses are fibrous joints that hold some long bones together such as the tibiofibular syndesmosis. They allow a small degree of mobility. Gomphoses are immobile fibrous joints found only between the teeth and their bony sockets.
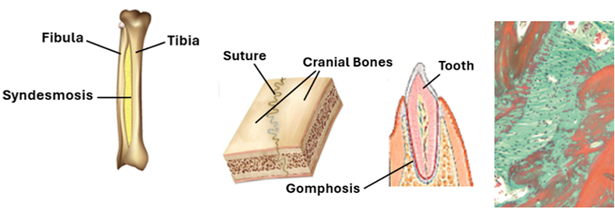
Cartilaginous Joints
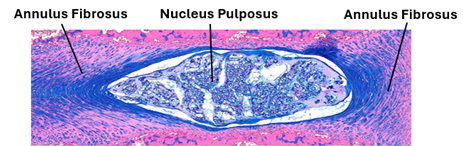
In this type of joints bones are joined together by cartilage. Cartilaginous joints are of types of primary cartilaginous joints made of hyaline cartilage and secondary cartilaginous joints that contain fibrocartilage. The primary cartilaginous joints are also known as synchondroses; they are made of hyaline cartilage. In adults, synchondrosis (primary cartilaginous joints) are present in the first sternocostal and between the 11th rib and the costal cartilage. They do not allow movement of articulated bones. Secondary cartilaginous joints are also known as symphyses; they are made of fibrocartilage. They are strong joints that could allow limited amounts of movement (narrow symphysis) as in the pubic symphysis, or a wider range of movement (wide symphysis) as in the intervertebral discs. Intervertebral discs a central gelatinous part called the nucleus pulposus surrounded a concentric ring of fibrocartilage forming the anulus fibrosus. They provide strong articulation with cushioning and allow movement of vertebrae.
Synovial Joints
Synovial joints are the most common type of joints in the human body. They are also known as diarthroses because they allow free movement of the articulating bones. They are characterized by the presence of a fluid filled cavity called the synovial cavity. The fluid is called the synovial fluid and is secreted by the synovial membrane which surrounds the cavity. In this type of joints, the ends of the articulating bones are covered by hyaline cartilage known as the articular cartilage. The synovial membrane is surrounded by the joint capsule and joint ligaments.
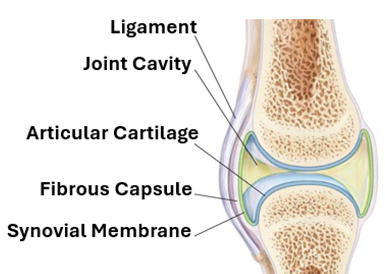
The Joint capsule
The joint capsule or articular capsule is a fibrous capsule that provides stability to synovial joints. It attached to the articulating bones and forms a sleeve around the joint. It consists of a dense irregular collagenous connective tissue that contains proprioceptive nerve endings. The inner part of joint capsule lining the joint cavity is called the synovial membrane.
The Synovial membrane
The synovial membrane, which is also known as the synovium, is a layer of loose connective tissue that lines the synovial cavity. It is often composed of two layers; a thin internal intimal layer and a deeper subintimal layer. The subintimal layer is made of loose areolar connective tissue. The intimal layer is a cellular layer that comprises two types of intimal cells called type A and type B synoviocytes. It produces and absorbs synovial fluid and participates in blood / synovial fluid exchange. Type A synoviocytes (type A synovial cells). Type A synoviocytes are phagocytic cells derived from blood monocytes. They are resident tissue macrophages that actively phagocytose cells debris and waste in joint cavity. The cytoplasm contains a well-developed Golgi, vesicles and lysosomes. Type 2 synoviocytes have an abundance of rER and well-developed Golgi complexes. They are characterized by extensive cytoplasmic thin processes passing in different directions, particularly towards the synovial cavity. Type 2 synoviocytes produce synovial fluid, collagen, mucin, lubricin and hyaluronic acid.
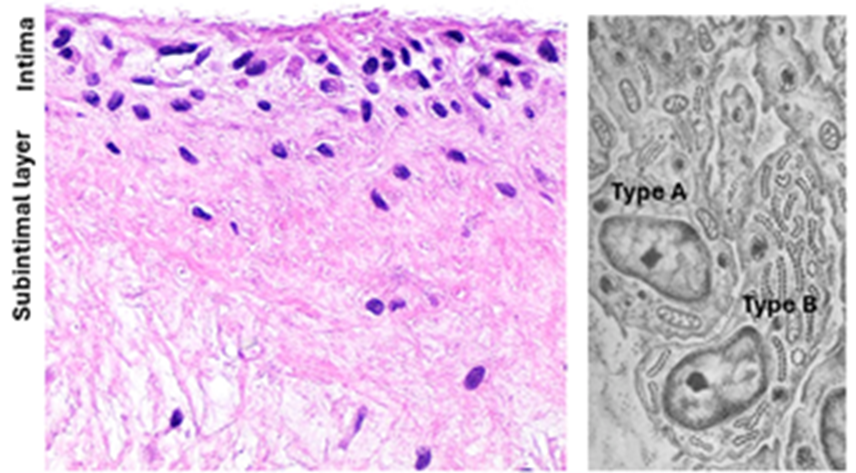
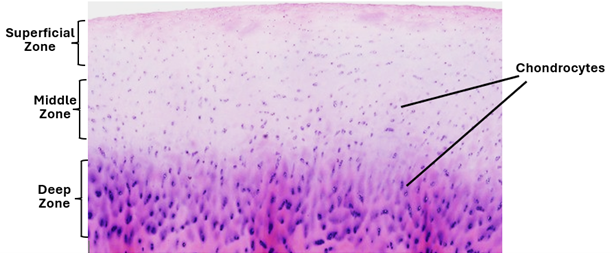
The articular cartilage
The articular cartilage completely covers the articular surfaces of the articulating bones. It provides smooth articular surfaces that facilitate friction free joint movement. It is about a plate of hyaline cartilage about 2-4mm in thickness. It has three zones; a superficial zone where chondrocytes are flattened and horizontally arranged, a middle zone widely separated spherical chondrocytes, and a deep zone where the chondrocytes are arranged in vertical columns. The surficial layer is not covered by a perichondrium, instead it is direct contact with the synovial fluid. It contains closely packed type 2 collagen fibers oriented parallel to the surface. It is resistant to shear and compressive forces. The middle zone constitutes about 50% of the size of the articular cartilage. It contains thicker collagen fibers that are oriented obliquely. In the deep zone, collagen fibers are lager and arranged perpendicular to the basal surface, providing the highest degree of resistance to compressive forces. The matrix of the articular cartilage is basophilic and amorphous and may show territorial and interterritorial regions. It contains water, collagen fibers, proteoglycans, inorganic ions, and glycoproteins. The vast majority (90-95%) of collagen is types 2 collagen; other forms of collagen including type 1 collagen are sparse. Matrix components are secreted and maintained by chondrocytes.
The articular cartilage is devoid of blood vessels, nerves and lymphatics. It has a limited regenerative capability. Nutrition of the articular cartilage is accomplished by diffusion from the synovial fluid.
The synovial fluid
Synovial fluid is a viscid physiologic fluid that fills the synovial cavity. It acts as a lubricant to the articular cartilage and source of nutrition to the articular cartilage. It is an ultrafiltrate of blood plasma that contains macromolecules produced by type B synoviocytes. The macromolecules produced by type 2 synoviocytes include hyaluronic acid, lubricin and proteinases.

Comments