
Muscular Tissue
- teachanatomy

- Apr 12
- 18 min read
Muscle Tissue constitutes about 40% of the human body weight. It is one of the four primary tissues of the body. Muscle tissue is responsible for movement of the body (locomotion), or body organs (e.g. intestinal peristalsis) or movement of parts of the body or its organs. It does so by contracting; contraction is a characteristic feature of muscle cells which constitute muscle tissue. Muscle cells are often referred to as muscle fibers and are usually closely packed. Structurally muscle tissue may show cross striations and are known as striated muscle cells/fibers or are devoid of cross striations and are known as non-striated muscle fibers. Functionally, muscle fibers could be under the control of the will (voluntary) or out of control of the will (involuntary). It is of three types
1. Skeletal muscle
2. Cardiac muscle
3. Smooth muscle

Skeletal Muscle:
Skeletal muscle forms the body’s muscle mass. It is voluntary and shows cross striations under the microscope. Skeletal muscles contain huge numbers of skeletal muscle fibers; large muscles containing thousands of muscle fibers. Within each muscle the fibers are arranged in bundle known as fascicles. Fascicles could be large or small depending on the size of the muscle itself. Large muscles with large fascicles have coarse textures. Fascicles may not usually extend the full length of the muscle, also fibers may or may not extend over the entire length of the fascicle.

Skeletal fibers are huge cells. They are the largest cells of the human body. A skeletal muscle cell (fiber) may be up to 40 cm long and up to 100um thick. It is cylindrical and multinucleated containing several to a couple of hundred nuclei. Each fiber (cell) is packed with small cylindrical structures known as myofibrils. In turn, myofibrils are packed with thread-like protein structures known as myofilaments. The myofilaments are of two types, myosin filaments and actin filaments. Myosin filaments are thicker than actin filaments. Actin myofilaments and myosin myofilament are regularly arranged within myofilaments which in turn are well organized within the muscle cell so that the myofibrils and muscle cell (fiber) itself show distinct regular cross striations of alternating light and dark bands.
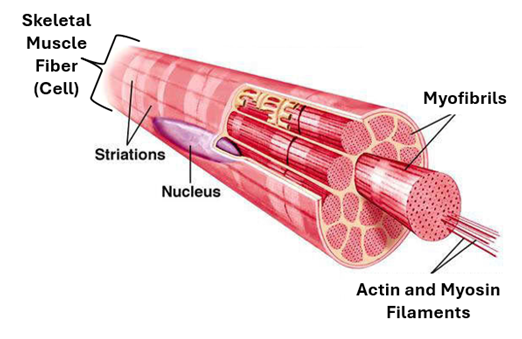
The Skeletal Muscle Fiber: This is the structural and functional unit of skeletal muscle. It is quite a long thick multinucleated cell that is It is usually cylindrical with tapering ends; its ends may occasionally bifurcate. In H&E-stained histological sections, the skeletal muscle fiver appears deeply acidophilic, due to high content of proteins (actin and myosin). The skeletal muscle fiber (cell) is covered by a specialized cell membrane called sarcolemma. The diameter of skeletal muscle fibers increases with age and exercise. Their size also increases in response to the male sex hormone (testosterone). The nuclei are numerous, (about 35 nuclei/mm of fiber length) and are peripherally located.

The nuclei are elongate, oval, basophilic nuclei and are arranged parallel to the surface of the fiber. The myofibrils are arranged parallel to each other and to the long axis of the fiber. They extend over the entire length of the fiber and do not branch. They are 1-2 um in diameter and show regular cross striations. Cross striations of all fibrils are exactly aligned to each other, imparting same cross striations on the muscle fiber

The sarcoplasm is the term given to the cytoplasm of muscle cells (fibers). The prefix sarco- meaning “belonging to muscle or flesh”. Since the interior of the skeletal muscle cell is almost fully occupied by myofibrils, the sarcoplasm is confined to the peripheral regions of the muscle fiber around the nuclei and also in the spaces present in between the myofibrils. The sarcoplasm contains a variety of cytoplasmic organelles, including mitochondria, Golgi apparatus, lysosomes, ribosomes and smooth endoplasmic reticulum (sER). sER of muscle fibers (cells) is known as sarcoplasmic reticulum. The sarcoplasm also contains inclusions, which include myoglobin, glycogen particles, lipid droplets and lipofuscin. There are two types of muscle fibers red and white muscle fibers. Mitochondria and myoglobin are more abundant in red muscle fibers than in white muscle fibers. The sarcoplasm also contains troponin and intermediate filaments, such as desmin, nestin and vimentin. It also contains troponin which plays an important role in muscle contraction when activated by calcium.
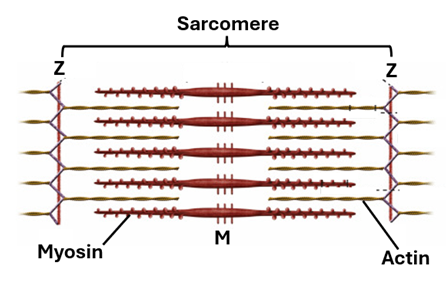
Striation is a characteristic feature of skeletal muscle fibers. It is the presence of alternating light and dark bands. The light bands are known as the I-bands, whereas the dark bands are known as the A-bands. When examined with polarized light microscope, skeletal muscle fibers show light bands where light is not refracted, they are isotropic (I = isotropic). On the other hand, there are dark bands where light is refracted; these are the anisotropic bands (A= anisotropic). In the middle of the I-band there is a thin dark line called the Z line. Also, in the middle of the A-band, there is a light narrow band called the H-band. In the middle of the H-band, there is a line known as the M-line. That part of the myofibril between any two successive Z-line is the sarcomere. It is the contraction unit of striated muscle. Electron microscopy reveals that I-bands contain actin filaments only, whereas A-bands contain both myosin and actin filaments. The H-band contains myosin filament only. The width of the H-band is variable. When the muscle is relaxed the H-band is wide, and when it is contracted the H-band is narrower. This is because muscle contraction takes place due to gliding of actin filament in between the myosin filaments towards the M-line.

Banding under the Microscope. Skeletal muscle banding is clearly visible under both the light and electron microscope. Under the light microscope the dark and light bands (A-, I- & H-bands) are clearly visible with medium power magnification (X40). Under the electron microscope, all three bands along with Z and M line are clearly visible. Moreover, actin filaments, myosin filaments, mitochondria, sarcoplasmic reticulum, T-tubules and other contents of the sarcoplasm are visible under the electron microscope.

The Sarcoplasmic Reticulum. Muscle fibers (cells) are rich in smooth endoplasmic reticulum (sER). sER of muscle fibers is unique and is known as the sarcoplasmic reticulum. It is complex and plays an important role in muscle contraction; it stores calcium ions and releases them when the muscle fiber is stimulated by a nerve impulse. The sarcoplasmic reticulum encircles myofibrils, arranging them into bundles known as Koelliker’s columns. The sarcoplasmic reticulum itself is arranged into sets. Each set consists of two terminal cisternae and many tubules in-between. Cisternae of adjacent sets are associated with a transverse (T-) tubule. The T-tubule is an invagination of the sarcolemma (cell membrane of the muscle fiber). The two cisternae and T-tubule constitute what is known as the triad. In skeletal muscle, the triad encircles the myofibril at the level of the A-I junction.
The transverse tubule (T-tubule) is a tubular structure formed by invagination of the sarcolemma (cell membrane) into the interior of the striated muscle fiber. Several T-tubules traverse the muscle fibers at intervals. They accompany cisternae of sarcoplasmic reticulum as they wrap bundles of myofibrils. Each T-tubule is flanked on either side by a terminal cisterna of sarcoplasmic reticulum, the three together forming a triplet known as the triad. The T-tubule is an extension of the cell membrane. Thus, when the plasmalemma (sarcolemma) of the skeletal muscle fiber is excited by a motor nerve ending the wave excitation is carried along the T-tubule into the interior of the nerve fiber. This wave stimulates the terminal cisterns of sarcoplasmic reticulum which in response release calcium ions into the sarcoplasm facilitating gliding of actin filament in between myosin filaments causing sarcomere shortening (muscle contraction0

Actin, Myosin and Muscle Contraction. Muscle contraction takes place by gliding of actin filaments between myosin filaments in the presence of calcium ions and adenosine triphosphate (ATP). ATP provides energy required for contraction and calcium facilitates binding of actin filaments to myosin filaments. Myosin filaments have actin binding sites, but normally these are by tropomyosin. Troponin binds to calcium ions exposing the myosin binding sites. Once these sites are exposed actin filaments bind to myosin filaments and cross-bridge cycling begins. Actin filaments are pulled towards the M-line, the I-bands and H-band become narrower, and the sarcomere shortens causing muscle fiber contraction.
Connective Tissue Elements of Skeletal Muscle: The connective tissue contributes to muscle functioning and organization. The muscle as whole is surrounded by a tough fibrous connective tissue sheath known as epimysium, which encompasses fascicles of skeletal muscle fibers. Each fascicle is surrounded by a fibrous connective tissue capsule known as perimysium. Within each fascicle, muscle fibers are embedded in a delicate loose connective called endomysium. Endomysium is made of collagen type 1 fibers, elastic fibers and reticular fibers, in addition to the ground substance, fibroblasts, blood vessels, lymphatic vessels and nerve fibers.
Epimysium is a tough dense irregular fibrous connective tissue that envelops the whole muscle protecting it and reducing friction against surrounding bones and other tissues. It helps the muscle to maintain its integrity. It is continuous with the perimysium and with tendons thus conduction the power of muscle contraction to bones facilitating locomotion. Large blood vessels and nerves gain access to the muscle via the epimysium to the perimysium to the endomysium to reach the individual muscle fibers. The endomysium wraps the individual muscle fibers and separates them from each other. It allows friction free movements of the individual fibers. It brings blood vessels and nerve fibers to the muscle fibers.

Innervation of the Skeletal Muscle: Skeletal muscle is very well innervated. Each skeletal muscle fiber has its own motor innervation. Motor (efferent) nerve fibers end on muscle fibers forming motor endplates. As it approaches the muscle fibers, the efferent nerve fiber gives off several branches. Each of these branches innervates a single muscle fiber (making synaptic junctions). The nerve endings are clearly enlarged terminals of the nerve fibers. They contain a few mitochondria and numerous small clear vesicles full of the neurotransmitter acetylcholine and are thus called cholinergic vesicles and the ending is designated as cholinergic. The neurilemma of the ending makes synaptic junction with the sarcolemma. Here, both membranes thicken; that of the muscle fiber (sarcolemma) is thicker and is known as the postsynaptic membrane. Cholinergic vesicle fuse with the presynaptic membrane of the neurilemma and pass out their content (acetylcholine) into the synaptic cleft (the space between the neurilemma and the sarcolemma). Acetylcholine stimulates the sarcolemma. The wave of excitation is carried along the sarcolemma into the depth of the muscle fiber via the T-tubules. In response to the wave of excitation in the T-tubules, the sarcoplasmic reticulum releases its content of calcium thus initiating gliding of actin filament between myosin filaments causing contraction of the muscle fiber.

Sensory Innervation: All skeletal muscle fibers receive motor innervation but only a few have sensory innervation. These muscle fibers and their sensory endings constitute muscle spindles. The Muscle spindle is a group of small skeletal muscle fibers surrounded by a capsule, and assuming a spindle shape. The fibers within the spindle are known as intrafusal; they are of two of types; nuclear bag fibers and nuclear chain fibers. In the nuclear chain fiber, the nuclei are arranged in a row, whereas in nuclear bag fibers they are grouped into a spherical mass. Both nuclear bag and nuclear chain fibers sense stretch, but nuclear chain fibers sense sustained stretch. Both are innervated by mechanoreceptor endings that sense stretch and deformation. The sensory nerve endings surround the spindle muscle fibers either spirally or in a flower spray fashion.

Cardiac Muscle:
Cardiac muscle is striated and involuntary. It consists of branching and anastomosing muscle fibers. Cardiac muscle fibers together with each other form a structural and functional syncytium. Unlike the skeletal muscle fiber, which is a single multinucleated cell, the cardiac muscle fiber is not a single cell, but a group of muscles cells known as cardiac myocytes attached to each other end-to-end. Thus, whereas skeletal muscle fiber and skeletal muscle cell stand for the same structure, the cardiac muscle fiber and the cardiac muscle cell are two different structures. Cardiac muscle fibers are long branching cylindrical structures made of cardiac muscle cells. They branch and anastomose with each other. Cardiac muscle cells (cardiac myocytes) are components of cardiac muscle fibers. Within the cardiac muscle fiber, cardiac muscle cells are joined end to end and held together by intercalated discs, which are a special type of cell junctions. Thus, intercalated discs delineate individual cardiac myocytes. The cardiac myocyte is a short branching cylindrical structure about 100um long and about 15um in diameter. Cardiac myocytes contain a single oval centrally located nucleus; two nuclei are occasionally seen.

As the case is with skeletal muscle fibers (cells), cardiac myocytes contain myofibrils which are packed with regularly arranged actin filaments and myosin filaments. Thus, they contain A-bands, I-bands, H-bands, Z-lines and M-lines. As in skeletal muscle that part of the myofibril between two successive Z-lines is known as the sarcomeres. The sarcoplasm of cardiac myocytes contains numerous long mitochondria, lipid droplets and glycogen particles. In the perinuclear region, the sarcoplasm also cytoplasm contains Golgi complexes and lipofuscin granules.
As is skeletal muscle, cardiac muscle cells contain transverse tubules (T-tubules) which are invaginations of the sarcolemma. In cardiac muscle, the T-tubules are present at the level Z-lines. This is different from the situation in skeletal muscle where the T-tubules are present at the level of the A/I junction. T-tubules function in carrying the wave of electrical impulses to cisternae of sER (sarcoplasmic reticulum) in deep parts of the myocytes.
Cardiac muscle is well vascularized. Individual muscle fibers are invested by a loose connective tissue equivalent to the endomysium. This endomysial loose connective tissue is highly vascular. It is quite rich in capillaries, that form networks around individual muscle fibers. This is to satisfy the high need of cardiac myocytes for oxygen and energy.
Cardiac muscle does have the capability to renew itself because cardiac myocytes are fully differentiated cells incapable of going back into the cell cycle, and they do not have progenitor cells that would differentiate into functional myocytes to replace lost myocytes. Thus, if cardiac myocytes are lost due to deprivation from the blood supply (infarction) or any other pathology, they are not replaced by new myocytes but by a fibrous connective tissue. The thus formed fibrous connective tissue interrupts the spread of the wave of excitation across cardiac muscle, thus interfering with the normal process of muscle contraction, and leads to malfunctioning of the heart.

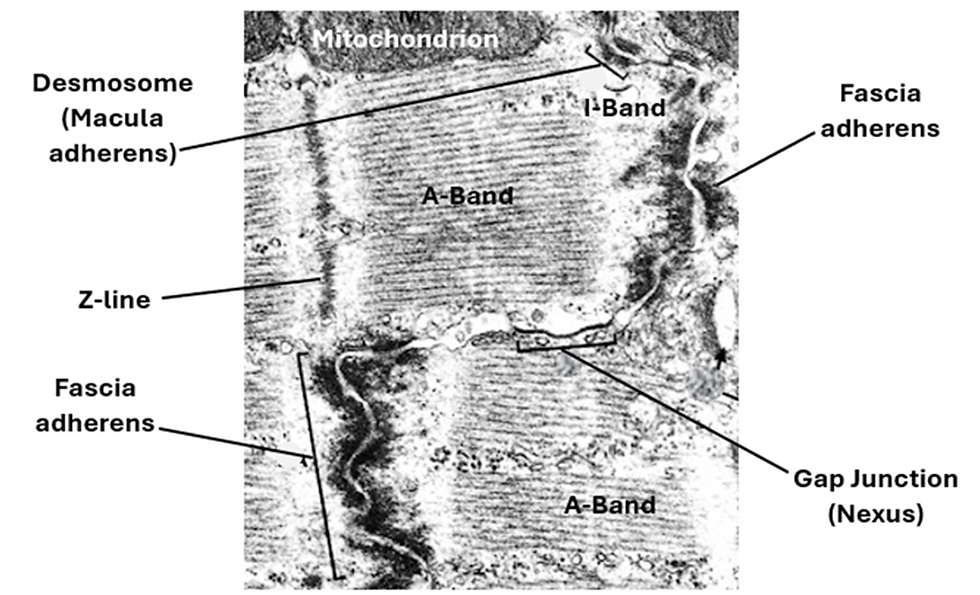
Intercalated discs (ID): Intercalated discs are a characteristic feature of cardiac muscle. They are specialized mechanical and electrical intercellular junctions. They are seen by both the light and electron microscope. In H&E sections under the light microscope they appear as transverse lines resembling the cross striations and running parallel to them but are slightly thicker. They are better seen in histological sections stained by special techniques (such as trichrome method and immunohistochemical methods).

Under the electron microscope, intercalated discs show a step-like appearance, each consisting transverse and lateral components. Transverse components run parallel to the Z-lines whereas the lateral components run parallel to the action and myosin filaments. Intercalated discs comprise three kinds of cell junctions. These are fascia adherents, desmosomes and gap junctions. Fascia adherents are long wavy ribbon-like cell junctions. They resemble the zonula adherents of epithelial membranes. Fascia adherents are found in the transverse component of the intercalated disc and serve as anchorage sites for actin filaments of adjacent myocytes. Desmosomes are also known as macula adherents. Are disc- or button-shaped cells junction They are found in both the transverse and lateral components of the intercalated disc. They hold together adjacent myocytes. Both fascia adherents and desmosomes are characterized by obvious thickening of the cell membranes (sarcolemma) of the adjacent cells. Gap junctions are also called nexuses. Here, the cell membranes (sarcolemma) of adjacent cells come closer to each other to be separated by a narrow gap only 2nm wide. This narrow gap is traversed by channels that allow exchange of ions between the adjacent myocytes. Gap junctions are found in the lateral components of the intercalated discs. Gap junctions are sites where the electrical impulses spreading along the sarcolemma pass from one myocyte to the adjacent one. Thus, gap junctions allow conduction of the contractile impulse from one myocyte to another. They facilitate electrical communication between the myocytes thus constituting a functional syncytium. As such, the intercalated discs join adjacent cardiac myocytes together both mechanically and electrically.
Modified Cardiac Muscle Cells:
In certain parts of the heart, the cardiac muscle cells (cardiac myocytes) are modified to carry out specific functions. These include the secretory myocytes, Purkinje fibers and atrial nodal cells.
Secretory Myocytes: Secretory myocytes are few but are present in both the atria and ventricles. Atrial myocytes synthesize atrial natriuretic peptide (ANP) and store it in specific dense granules (SG). Atrial granules are numerous in the perinuclear region. When atrial myocytes are overstretched, in the case of heart failure for instance. In response to stretching, the atrial myocytes release ANP by exocytosis. ANP is a hormone that reduces extracellular fluids by increasing glomerular filtration of Na+ and reduction of its absorption. Ventricular myocytes produce brain natriuretic peptide (BNP). BNP has a paracrine function. It acts locally to reduce ventricular fibrosis.

Purkinje Fibers. Purkinje fibers are modified cardiac myocytes specialized in impulse conduction. They are large spindle-shaped cells with a single nucleus. They contain fewer myofibrils than myocytes. Glycogen is present in abundance around the nucleus; thus, the perinuclear region appears pale (glycogen is water soluble and is washed away during tissue preparation. Purkinje fibers make up the bundle of His, and are part of the conducting system of the heart. They have desmosomes (anchorage) and gap junctions (impulse conduction)

The Nodal Cells: The nodal cells are fusiform and are smaller and paler than the surrounding myocytes. They lack intercalated discs. They are present in two groups forming the sinoatrial (SA) node and the atrioventricular (AV) node. The SA node is a small (about 25x5x2mm) group of nodal cells present in the upper part of the right atrium close to the orifice of the vena cava. It often surrounds a small artery known as SA node artery. Cells of the SA node are capable of spontaneous production of rhythmic electrical impulses (action potential) and thus they set the pace for contraction of the heart muscle. Accordingly, the SA node is known as the Pacemaker of the heart. The SA node is innervated by sympathetic and parasympathetic nerve fibers, which influence the firing frequency of SA nodal cells increasing or decreasing heart rate.
AV Node
The AV node is present in the lower back region of the interatrial septum. The AV nodal cells are modified cardiac myocytes with many branching processes. They are paler than the ordinary cardiac myocytes. Impulses from SA node travel in the atrial walls through the atrial fibers until they reach cells of the AV node. AV nodal cells receive the impulse and delay it prior to its transmission via the bundle of His.

Smooth Muscle:
As a tissue, smooth muscle is well distributed in the human body. It develops from the mesoderm and the neural crest. Smooth muscle is found in the walls of hollow visceral organs such as the intestines, stomach, parts of the esophagus, the trachea, bronchi, bronchioles, the ureter, the bladder, and in the walls of blood vessels. It is found in many other locations such as the dermis of the skin, ciliary body of the eye. Wherever it is present it contributes to the normal functioning of those organs and structures. It participates in the process of food digestion and its movement within the alimentary tracts and in regulation of blood pressure. Smooth muscle is involuntary and nonstriated. It is not under the control of the person’s will but under control of the autonomic nervous system. It carries on its work without interference from the person. It is made of spindle-shamed cells. Smooth muscle is non-striated and involuntary. Smooth muscle is not as strong as skeletal and cardiac muscle but has the capability of maintaining contractions for long periods of time.
Smooth muscle is made of fusiform or spindle-shaped cells that lack cross striations. The smooth muscle cell is often called a smooth muscle fiber. Each fiber (cell) is 50-200um in length and 2-5um in diameter and has a single oval nucleus. The nucleus contains one or two prominent nuclei. Smooth muscle cells contain actin and myosin filaments that are not regularly arranged. There are no H- and I-bands nor Z-lines, nor sarcomeres. Smooth muscle cells contain sarcoplasmic reticulum which controls calcium ion flow as the case is in skeletal and cardiac muscle. The sarcoplasm of smooth muscle fibers contains many dense bodies that act as sites of anchorage for filaments. Smooth muscle cells do not contain troponin. Smooth muscle cells possess cell junctions which include desmosomes for anchorage and gap junctions for passage of impulses.
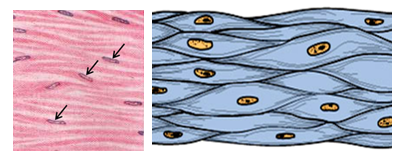
Motor Innervation of Smooth Muscle: Based on the mode of motor innervation, smooth muscle is classified into two types: single unit smooth muscle and multiunit smooth muscle. In the single unit smooth muscle, a single nerve ending induces the contraction of numerous smooth muscle fibers. This type of smooth muscle is characterized by the presence of intercellular gap-junctions between a group of smooth muscle fibers. The nerve ending makes synaptic junctions with only one muscle fiber (cell). Only this cell is excited. Then, a wave of excitation spreads along the sarcolemma of cell (fiber) to the neighbouring ones by passing from one cell to the other across the intercellular gap-junction, specifically across connexin channels of the gap junctions. Thus, contraction of the muscle fibers occurs synchronously. Single unit smooth muscle is found in locations like the alimentary tract where synchronized contraction of many smooth muscle fibers is needed for carrying on the specific functions of the organ such as peristalsis. The multiunit smooth muscle on the other hand, lacks gap junctions. Not only that, but also each of its smooth muscle fibers has its own motor innervation and its own neuromuscular synaptic junction. Thus, contraction of the fibers in this type of muscle is not synchronized; each muscle fiber contracts on its own. Multiunit smooth muscle is found in locations where precision contraction is needed such as smooth muscle of the iris and ciliary body of the eye, and also large arteries and respiratory passages.


Comments